Marine nutrient cycling - How will the ocean’s capacity of biological carbon pumping change? [Present]
Christiane Lancelot
Ecologie des Systèmes Aquatiques, Université Libre de Bruxelles, Belgium; lancelot@ulb.ac.be
The biological pump refers to a suite of biologically mediated processes that transport carbon from the ocean’s surface layer to its interior. Its efficiency depends on the balance between the rates of carbon photo-assimilation, export and mineralization. Our knowledge of the biological carbon pump relies on our mechanistic understanding of factors structuring phytoplankton distributions and marine food webs, and the associated biogeochemical cycles. To assess the extent to which the strength and the efficiency of this pump will change in the future, we need to know how these factors – light, nutrients and temperature – might change in a warmer ocean. Models coupling an ecosystem module to a global circulation model provide important tools for understanding the dynamics of the carbon pump and its response to warming. But as pointed out by Sarmiento et al. (2004), existing tools are still not mature enough to allow this.
The last decade has seen increasing awareness of the relationship between key phytoplankton groups and their pivotal role in the functioning of the biological carbon pump (e.g. Boyd et al. 2010). Until recently, the picture was of a simple subdivision between efficient carbon export via a diatom-copepod-fish linear food chain in nutrient-rich waters and retention of surface carbon in nutrient-poor waters via a microbial network initiated by the ubiquitous pico/nano phytoplankton (Chisholm 2000). This picture has been recently complicated by the recognition of two additional small-sized players – the calcifying coccolithophores and the nitrogen-fixing cyanobacteria. These have a competitive advantage over diatoms in warm, well-illuminated surface waters supplied with imbalanced inorganic nitrogen and phosphorus nutrients. Their participation in carbon export is indirect and involves either aggregation with calcium carbonate liths acting as ballast particles or the release of nitrogen that sustains the growth of concomitant diatoms, thereby triggering carbon export (Chen et al. 2011).
 |
|
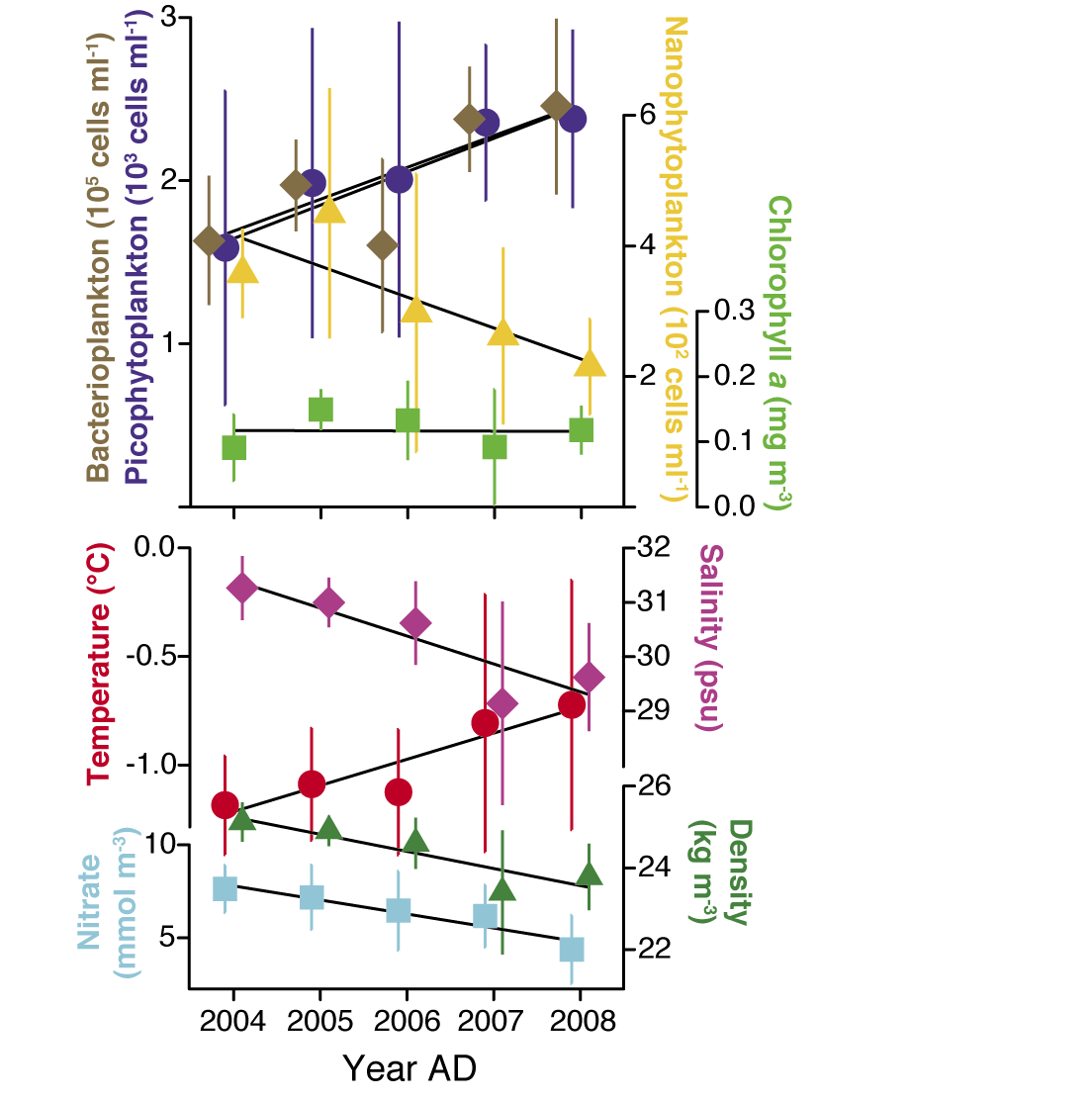
Figure 1: Summer conditions in the upper water layers of the Canadian Basin. The lower panel shows the physical water properties over the period 2004-2008, the upper the response of the plankton organisms during the same period. Figure modified from Li et al. 2009. |
Ocean warming affects the pelagic ecosystem both directly and indirectly, by increasing temperature and stratification. The latter tend to favor the dominance of small phytoplankton (e.g. Falkowski and Oliver 2007; Li et al. 2009) over large cells such as diatoms (Fig. 1). The resulting photo-assimilated carbon benefits the heterotrophic microbial food web, whose activity is stimulated by the warmer temperature (Sarmento et al. 2010), increasing the rate of carbon mineralization. Small phytoplankton cells also have low sinking rates. The spreading of such cells anticipated with increased ocean stratification will decrease the overall capacity of the biological pump, but the extent remains uncertain (Barber 2007).
The prevalent view of the picophytoplankton carbon being totally remineralized in the surface waters has been recently challenged by data from the equatorial Pacific Ocean and Arabian Sea, which point to significant export of picophytoplankton-related carbon through indirect paths such as aggregation and fecal pellets (Richardson and Jackson 2007). The future latitudinal extent of coccolithophore blooms due to warming is unknown, however, particularly in light of the possible alteration of calcification rates by ocean acidification (Cermeño et al. 2008).
The potentially large ocean deoxygenation due to the increased temperature and stratification projected for a warmer ocean (Keeling et al. 2010) will have direct consequences for marine biota, but only an indirect effect on ocean productivity and nutrient and carbon cycling. An expansion of suboxic/anoxic conditions would increase the release of phosphate and iron from sediments while some reactive nitrogen would be eliminated by denitrification or anaerobic ammonia oxidation. The subsequent shift in the ocean nitrate-to-phosphate balance will affect the composition and productivity of marine organisms, notably diazotrophic cyanobacteria, with uncertain consequences for the efficiency of the biological pump.
On the whole, the smallest phytoplanktons seem to have a competitive advantage in a warmer ocean. In contrast, diatoms are at an advantage in surface waters with transient nutrient pulses. In particular, they will benefit in coastal regions from stronger wind-driven upwelling events that are expected from increased storm and frequency resulting from climate warming. Improving the evaluation of changes to the biological carbon pump via ocean models is currently hampered by several uncertainties on mechanisms controlling phytoplankton dominance and food-web structures. Also, many global circulation models remain coarse in resolution and don’t serve high frequency forcing. To be able to predict better the efficiency of the pump under future conditions, the complexity in biology needs to be matched with an appropriate complexity in the representation of the physical and chemical environment in ocean models.
references
Full reference list online under: http://pastglobalchanges.org/products/newsletters/ref2012_1.pdf
Boyd PW, Strzepek R, Fu F and Hutchins D (2010) Limnology and Oceanography 55(3): 1353-1376
Chen Y-L L, Tuo S-H and Chen H-Y (2011) Marine Ecology Process Series 421: 25-38
Keeling RF, Kortzinger A and Gruber N (2010) Annual Reviews of Marine Science 2: 199-229
Richardson TL and Jackson GA (2007) Science 315: 838-840
Sarmento H et al. (2010) Philosophical Transactions of the Royal Society B 365: 2137-2149